


El citoplasma celular se encuentra delimitado por la membrana celular y por la membrana nuclear, y contiene a los diversos orgánulos celulares.
Los orgánulos celulares pueden clasificarse atendiendo a su posesión de membrana, y según sea una membrana simple o doble, en:
- Los orgánulos sin membrana, como son los ribosomas e inclusiones citoplasmáticas.
- Orgánulos con membrana simple: retículo endoplasmático, aparato de Golgi, lisosomas, peroxisomas y vacuolas.
- Los orgánulos con membrana doble, las mitocondrias y cloroplastos.
Orgánulos celulares sin membrana
En las células podemos encontrar orgánulos sin membrana. Estos orgánulos celulares son los ribosomas y las inclusiones citoplasmáticas.
Ribosomas
Los ribosomas son orgánulos celulares que no poseen membrana. Están formados por ARN y proteínas.
Los ribosomas están presentes en todo tipo de células, excepto en espermatozoides maduros. Se localizan libres en el citoplasma, en forma de polirribosomas, asociados al retículo endoplasmático rugoso, en su cara externa, o asociados a la membrana nuclear. Los polirribosomas son agrupaciones de ribosomas asociados a ARN mensajero.
La función de los ribosomas es la de participar en la síntesis de proteínas.
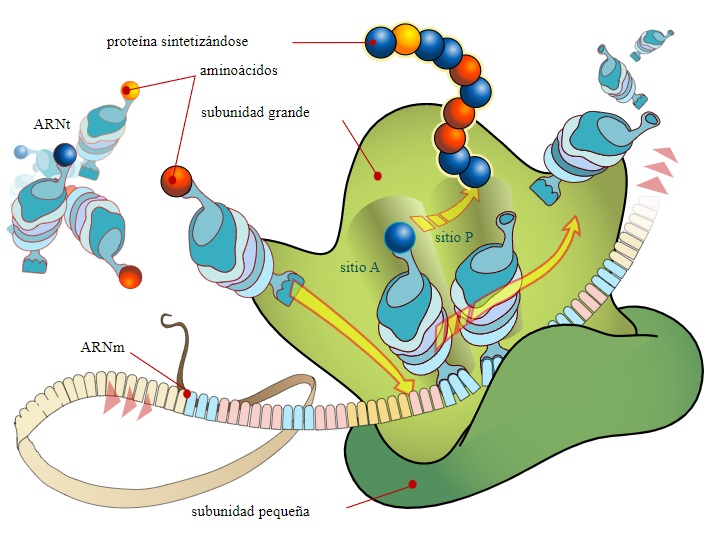
Tienen dos subunidades, la subunidad mayor y la subunidad menor. La subunidad mayor está formada por dos o tres moléculas de ARN y por proteínas. En cambio, la subunidad menor solo tiene un tipo de ARN asociado a proteínas. Las subunidades forman un surco, donde se asociará la proteína que se esté sintetizando. Y también un segundo surco, donde se aloja el ARN mensajero. Las subunidades del ribosoma se encuentran separadas en el citoplasma y sólo se unen cuando se va a proceder a la traducción (lectura del ARN mensajero).
El coeficiente de sedimentación es un indicador que informa sobre la velocidad con la que sedimenta una molécula o una partícula. Es un reflejo de su forma y su tamaño, y se mide en Svedbergs (1S = 10−13 seg.). En las células procariotas, en mitocondrias y cloroplastos, el coeficiente de sedimentación de los ribosomas es de 70S. En las células eucariotas, es de 80S.
Inclusiones citoplasmáticas
Las inclusiones citoplasmáticas son depósitos sin membrana en los que se acumulan sustancias de reserva o desecho, productos del metabolismo celular. Pueden ser gránulos de carbohidratos, gotas de lípidos…
Los gránulos de carbohidratos, son gránulos de glucógeno en células animales y de almidón en las vegetales. Son almacenes de energía.
Las gotas de lípidos abundan en semillas y frutos. En animales, las gotas de grasa se acumulan en el tejido adiposo, nuestra principal reserva energética.
Otras inclusiones pueden ser de látex, de pigmentos, proteínas cristalizadas, etc.
Orgánulos celulares con membrana simple
El retículo endoplasmático
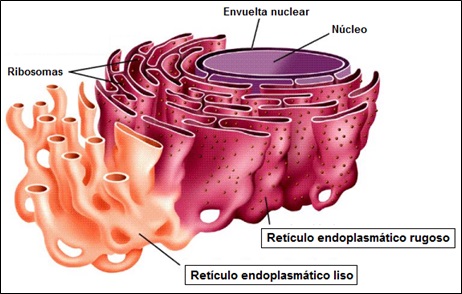
El retículo endoplasmático es un complejo sistema de membranas, compuesto por sáculos y túbulos aplanados conectados entre sí que delimitan un espacio interno o lumen. Los túbulos se comunican a su vez con otro orgánulo celular, el aparato de Golgi, además de con membrana nuclear externa. Existen dos tipos de retículo endoplasmático, el rugoso (RER) y liso (REL).
Retículo endoplasmático rugoso (RER)
El retículo endoplasmático rugoso está constituido por sistema de cisternas con ribosomas adheridos a la cara citoplasmática de su membrana, es decir, en su cara externa. Se encuentra presente en casi todas las eucariotas, excepto en los glóbulos rojos.
Tiene diversas funciones:
- Síntesis de proteínas, en los ribosomas de su cara externa.
- Modificación de proteínas o glucosidación, por transferencia de glúcidos, fundamentalmente oligosacáridos.
- Almacenamiento de proteínas. En el lumen se almacenan proteínas sintetizadas.
Retículo endoplasmático liso (REL)
El retículo endoplasmático liso está formado por túbulos ramificados intercomunicados entre sí y con el retículo endoplasmático rugoso. No contiene ribosomas asociados en ninguna de sus caras. Sus funciones son:
- Síntesis de lípidos de membrana. En la cara externa del REL se sintetizan prácticamente todos los lípidos de la célula, excepto los ácidos grasos y ciertos lípidos mitocondriales.
- Síntesis de hormonas derivadas del colesterol.
- Eliminación de la toxicidad de moléculas que resultan perjudiciales para la célula, como medicamentos, drogas, conservantes, insecticidas, etc, en células hepáticas.
- Almacenamiento de calcio, en células musculares. Libera el calcio como respuesta a estímulos nerviosos, para permitir la contracción muscular.
- Interviene en la producción de pigmentos.
Aparato de Golgi
El aparato de Golgi está formado por un conjunto de sacos aplanados con forma de disco rodeados de membrana (cisternas) y vesículas asociadas. Las cisternas se disponen formando pilas, los dictiosomas. Las células pueden contener uno o más dictiosomas dependiendo de cuál sea su función, Por ejemplo, las células secretoras del páncreas presentan aparatos de Golgi muy desarrollados. Las células que realizan actividad secretora suelen tener un solo dictiosoma.
Este orgánulo se encuentra en todas las células eucarióticas excepto en los glóbulos rojos de mamíferos.
Presenta polaridad. En los dictiosomas se diferencias dos caras con distinta estructura y función, la cara cis y la cara trans.
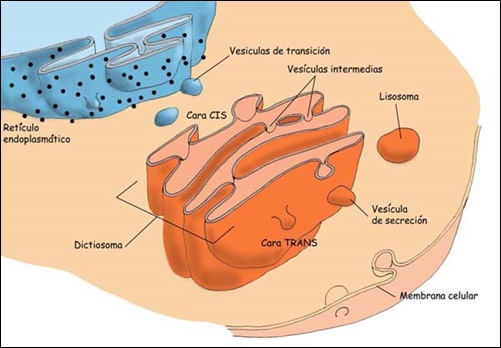
La cara de formación (cara cis) es la más próxima al núcleo de la célula. Se encuentra rodeada de pequeñas vesículas (vesículas de transición) que derivan del retículo endoplasmático. Esta cara está constituida por cisternas convexas conectadas con el retículo endoplasmático rugoso.
La cara de maduración (cara trans) es la que se encuentra más próxima a la membrana plasmática. De ella parten vesículas más grandes, vesículas secretoras.
Las sustancias procedentes del retículo endoplasmático llegan a la cara cis en forma de vesículas de transición. Estas vesículas de transición van pasando de cisterna en cisterna mediante vesículas de transporte (o intermedias), hasta que salen hacia los diferentes destinos celulares por la cara trans, en forma de vesículas secretoras.
Los destinos de las vesículas secretoras pueden ser dos. Se acumulan en el citoplasma originando lisosomas primarios, o se desplazan hasta la membrana plasmática y liberan su contenido por exocitosis.
Las funciones del aparato de Golgi son:
- En el aparato de Golgi se modifican, empaquetan, se transporta y se distribuyen moléculas sintetizadas en el retículo endoplasmático. Finalmente secreta estas moléculas.
- Regeneración de la membrana plasmática. La unión de las vesículas de secreción con la membrana produce la regeneración de esta última, pues se reponen los fragmentos perdidos por endocitosis.
- Glucosilación. Las proteínas y los lípidos completan la glucosilación que comenzó en el retículo endoplasmático mientras atraviesan las cisternas. Se eliminan y añaden nuevos azúcares, formándose las definitivas moléculas de glucolípidos y glucoproteínas.
- Este orgánulo participa además en la formación de pared celular en las células vegetales, y también en la formación del glucocalix en las células animales.
Lisosomas
Los lisosomas son pequeñas vesículas membranosas (~ 1mm) que contienen enzimas hidrolíticas implicadas en los procesos de digestión celular. Podemos encontrar dos tipos de lisosomas, los primarios y los secundarios.
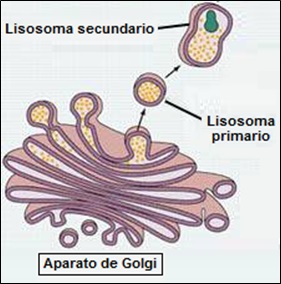
Los lisosomas primarios son los de reciente formación, procedentes del aparato de Golgi. Pueden verter su contenido al exterior por exocitosis o formar lisosomas secundarios.
Por otra parte, los lisosomas secundarios están formados por un lisosoma primario y una vacuola con material para digerir. Contienen, además de las enzimas hidrolíticas, sustancias en vías de degradación.
Entre los lisosomas secundarios, podemos distinguir fagolisosomas, que contienen material de origen exógeno. Es decir, procedentes de la nutrición celular y de la defensa ante los agentes patógenos. También podemos hablar de autofagolisosomas. Estos últimos contienen material de origen endógeno, destruyen materiales inservibles de la célula y aseguran la nutrición en condiciones desfavorables a partir de reservas.
Las enzimas hidrolíticas son las encargadas de catalizar las reacciones de hidrólisis. Este tipo de reacciones con aquellas en las que, mediante la intervención del agua, se rompen determinados enlaces covalentes: éster, peptídicos, glucosídicos, etc.). Tienen un pH óptimo próximo a 5 (hidrolasas ácidas), por lo que en la membrana de los lisosomas existe una bomba de H+ que, consumiendo energía (ATP), bombea iones hidrógeno desde el citosol de la célula con el objetivo de que en el interior del lisosoma se alcance el pH óptimo.
Estas enzimas deben estar confinadas dentro de los lisosomas por el peligro potencial que suponen para la célula. Si se encontrasen libres en el citoplasma, aún con el inconveniente de no estar en un medio donde el pH es el óptimo para su funcionamiento, podrían provocar la degradación de importantes estructuras celulares, causando la muerte de la célula.
Peroxisomas
Los peroxisomas son orgánulos de membrana sencilla que albergan en su interior enzimas oxidativos (peroxidasas y catalasas). Morfológicamente parecidos a los lisosomas.
Los enzimas oxidativos tienen como misión degradar sustancias que se forman como subproductos de algunas reacciones químicas y que pueden resultar perjudiciales para la célula. Como por ejemplo, el peróxido de hidrógeno, degradado a agua y oxígeno por el enzima catalasa.
Estos orgánulos se forman a partir del retículo endoplasmático rugoso por un proceso de gemación. Son vesículas carentes de ribosomas.
Los glioxisomas son un tipo especial de peroxisoma. Son exclusivos de las células vegetales (semillas en germinación), y contienen los enzimas necesarios para obtener azúcares a partir de ácidos grasos.
Otras funciones que realizan los peroxisomas son:
- Participan en la β-oxidación de los ácidos grasos en la célula.
- Intervienen en la síntesis de lípidos.
- Oxidan el exceso de NADH.
- Controlan los niveles de oxígeno.
Vacuolas
Las vacuolas son orgánulos citoplasmáticos rodeados de una membrana, originados a partir del aparato de Golgi, del retículo endoplasmático o de invaginaciones de la membrana plasmática. Se encuentran presentes en todo tipo de células, pero son especialmente abundantes en las células vegetales. Es posible encontrar en las células vegetales una única vacuola de gran tamaño, que ocupa entre el 30% y el 90 % del volumen celular.
Sus funciones son de almacén de sustancias, participan en la regulación osmótica de la célula y colaboran en la digestión celular. Son capaces de acumular en su interior sustancias hidrosolubles, como productos de desecho del metabolismo celular, sustancias de reserva, pigmentos, algunos alcaloides venenosos (opio, cianuro…), etc.
Su participación en la regulación de la presión osmótica se basa en el mantenimiento de la turgencia en células vegetales, pues acumulan el exceso de agua. En células animales destacan las vacuolas pulsátiles en protozoos que viven en medios hipotónicos, para bombear hacia el exterior agua.
Orgánulos con membrana doble
Mitocondrias
Las mitocondrias son orgánulos celulares que pueden tener diversa morfología (esféricos, cilíndricos, alargados…). Se encuentran presentes en todo tipo de células eucariotas. Estos orgánulos son especialmente abundantes en aquellas células con elevada demanda energética. En las mitocondrias se realiza la respiración celular, cuya finalidad es la de obtener la energía necesaria para la célula.
En su estructura se distingue una membrana mitocondrial externa y otra interna, el espacio intermembranoso y la matriz mitocondrial.
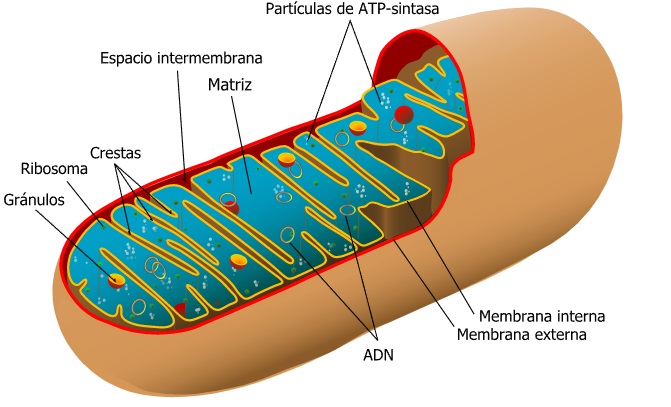
La membrana mitocondrial externa es muy permeable; contiene proteínas que forman canales. La membrana mitocondrial interna es más impermeable que la anterior. Esta última, la interna, presenta invaginaciones, llamadas crestas mitocondriales, que se introducen en la matriz. En ella se encuentran las cadenas de transporte electrónico y enzimas como la ATP-sintetasa.
El espacio intermembranoso es el espacio que existe entre ambas membranas mitocondriales, y se encuentra ocupado por una matriz semejante al citosol.
En la matriz mitocondrial existe ARN, ADN circular y ribosomas con coeficiente de sedimentación 70S. También incluye diversas enzimas, coenzimas, agua y sales minerales.
Como se ha mencionado, la función de las mitocondrias es la de la respiración celular. La respiración celular se basa en una serie de oxidaciones encaminadas a obtener energía en forma de ATP, entre otros. En la matriz de la mitocondria se realiza el ciclo de Krebs y la b-oxidación de los ácidos grasos. También se sintetizan proteínas mitocondriales a expensas de la maquinaria replicativa y del ADN mitocondrial. En la membrana mitocondrial interna se realiza el proceso de fosforilación oxidativa.
Cloroplastos
Los cloroplastos son también orgánulos celulares polimórficos. Captan la energía luminosa y fabrican, a través del proceso de fotosíntesis, biomoléculas necesarias para la vida de la célula. Se trata de un orgánulo característico de las células eucariotas vegetales.
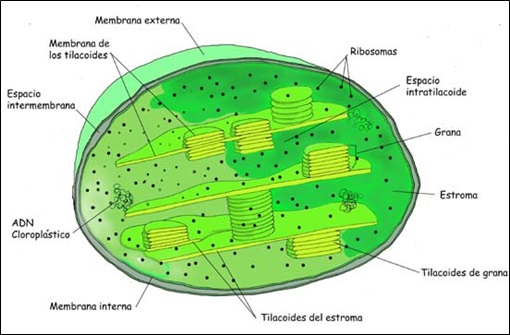
Podemos distinguir varias partes en su estructura. La membrana del cloroplasto, distinguiendo entre membrana externa e interna. La membrana externa es una membrana muy permeable. Sin embargo, la membrana interna es menos permeable que la anterior. Esta última posee proteínas específicas implicadas en el transporte. El espacio intermembranoso es el espacio que existe entre ambas membranas.
Los tilacoides son sáculos que se disponen paralelamente al eje mayor de los cloroplastos. Algunos se apilan formado los grana. Las membranas tilacoidales poseen pigmentos, como la clorofila. También podemos encontrar las cadenas de transporte electrónico y las ATP-sintetasas implicadas en el proceso de fotofosforilación.
El estroma es la matriz del cloroplasto. En el estroma existen moléculas de ADN circular, ribosomas con coeficiente de sedimentación 70S, gránulos de almidón y enzimas necesarias para la fijación del CO2.
Otros tipos de plastos
Además de los cloroplastos, en las células vegetales también podemos encontrar otros tipos de plastos, como pueden ser los cromoplastos y los leucoplastos.
Los cromoplastos son plastos que almacenan pigmentos tales como la clorofila o carotenos.
Por otra parte, los leucoplastos carecen de pigmentos. Se encuentran en los cotiledones, esbozos foliares del tallo y ciertas zonas de la raíz. Se trata de plastos que almacenan sustancias que pueden utilizarse como fuente de energía en los tejidos no fotosintéticos: los amiloplastos almacenan almidón, los oleoplastos almacenan aceites, y los proteinoplastos almacenan proteínas.
Más contenidos en www.loreseek.com
Síguenos en Twitter: @LoreSeekWeb y en Facebook: LoreSeekWeb
Estamos en Instagram: @loreseek




